Table of Contents
Glucose occurs naturally and is ubiquitous. It is one of the three most common monosaccharides; the other two are fructose and galactose. Monosaccharides are the most fundamental type of carbohydrates. They are called simple sugars as opposed to the more complex forms such as oligosaccharides and polysaccharides. They can combine, though, to form complex carbohydrates via glycosidic bonds (glycosidic linkages).
Glucose Definition
The term glucose is coined in 1838 by Jean Baptiste Dumas 1800 – 1884, a French chemist recognized largely for his works on organic analysis and synthesis. Etymologically, the term is based on the Ancient Greek γλεῦκος (gleûkos), meaning “sweetness”.
Biochemistry definition:
Glucose is an aldohexose monosaccharide (chemical formula: C6H12O6) that is ubiquitous in nature, and functions primarily as the main source of metabolic energy (the main substrate in glycolysis) in living things.
Etymology: Ancient Greek γλεῦκος (gleûkos, meaning “sweetness”). Abbreviation: Glu. IUPAC: (2R,3S,4R,5R)-2,3,4,5,6-Pentahydroxyhexanal. Chemical formula: C6H12O6. Synonyms: blood sugar; dextrose; corn sugar; d-Glucose; grape sugar.
Properties of Glucose
Glucose is a hexose monosaccharide. It is an organic compound. Its general chemical formula is C6H12O6. The molar mass of glucose is 180.156 g/mol. The melting point is 146 °C. Its heat capacity is 218.6 J K−1 mol−1. It is a crystalline solid, water-soluble, and sweet tasting.
Glucose vs. Galactose vs. Fructose
Glucose, galactose, and fructose are the three most common monosaccharides. Nevertheless, glucose is the most abundant. The three have the same chemical formula: C6H12O6. Hence, they are hexose monosaccharides, owing to the six carbon atoms. Both glucose and galactose are aldoses whereas fructose is ketose. Thus, glucose and galactose are more structurally alike. Nonetheless, glucose can be structurally identified from galactose based on the orientation of the hydroxyl group (OH) at carbon 4. Except for the hydroxyl group on the anomeric carbon (i.e. the carbon at which anomers rotate), the hydroxyl groups of glucose are in the equatorial position. Also, glucose has a lower melting point (146 °C vs. galactose’s 168–170 °C). It is also sweeter than galactose. However, of the three, fructose has the lowest melting point (i.e. 103 °C) and is the sweetest.
In terms of energy metabolism, glucose is the most commonly used since the organism readily utilizes glucose over galactose or fructose in glycolysis. In galactose metabolism, galactose enters glycolysis by first being phosphorylated via the enzyme galactokinase and then converted into glucose-6-phosphate. In fructose metabolism, fructose enters glycolysis by similarly going through certain steps prior to entering glycolysis. In humans, the conversion of fructose or galactose into glucose occurs primarily in the liver.
Types of Glucose
Two forms exist: Dextroglucose (D-Glucose) and Levoglucose (L-Glucose) based on Fischer projection. This nomenclature designates D– when the glucose stereoisomer rotates the plane-polarized light in the clockwise direction. L– is when it rotates the plane-polarized light in a counterclockwise direction. The naturally occurring and more common form is D-glucose. L-Glucose is produced synthetically.
Discovery of Glucose
Andreas Marggraf 1709 – 1782, a German chemist regarded as the pioneer of analytical chemistry, was credited for being the first to isolate glucose from raisins in 1747. He called it ‘eine Art Zucker’ (which means “a type of sugar”). As noted earlier, Jean Baptiste Dumas was cited as the one who called it glucose in 1838. Emil Fischer 1852 –1919, a German chemist and a recipient of the 1902 Nobel Prize in Chemistry, was attributed as the first to delineate the chemical structure of glucose. He was also known for establishing the stereochemical configuration of sugars, including glucose. Otto Fritz Meyerhof 1884 – 1951, a German physician cum biochemist and the recipient of the Nobel Prize in Physiology and Medicine in 1922, discovered glucose metabolism, particularly glycolysis, in 1922.
Common Biological Reactions
Below are some of the common biological reactions involving glucose.
-
Dehydration synthesis
Through dehydration synthesis, a monosaccharide, such as glucose, binds to another monosaccharide with the release of water and the subsequent formation of a glycosidic bond. The joining of two monosaccharides produces a disaccharide whereas the joining of three to ten monosaccharide units forms an oligosaccharide. Polysaccharides are produced by the joining of multiple monosaccharides. In this regard, glucose joins with another monosaccharide to form a disaccharide. For instance, two glucose molecules joined together by α(1→4) glycosidic bond form maltose, by α(1→1)α glycosidic bond form trehalose, and by β(1→4) glycosidic bond form cellobiose. Lactose forms when glucose and galactose combine, and sucrose, when glucose and fructose combine. Glucose is the major monosaccharide constituent of certain polymers like cellulose, starch, and glycogen. Polysaccharides made up of only glucose are called glucans.
-
Photosynthesis
Plants and other photosynthetic autotrophs produce their own food source (carbohydrates) through photosynthesis. The process uses carbon dioxide, water, inorganic salts, and light energy (e.g. from sunlight) captured by light-absorbing pigments, such as chlorophyll and other accessory pigments to produce hexose (e.g. glucose), water, and oxygen molecules.
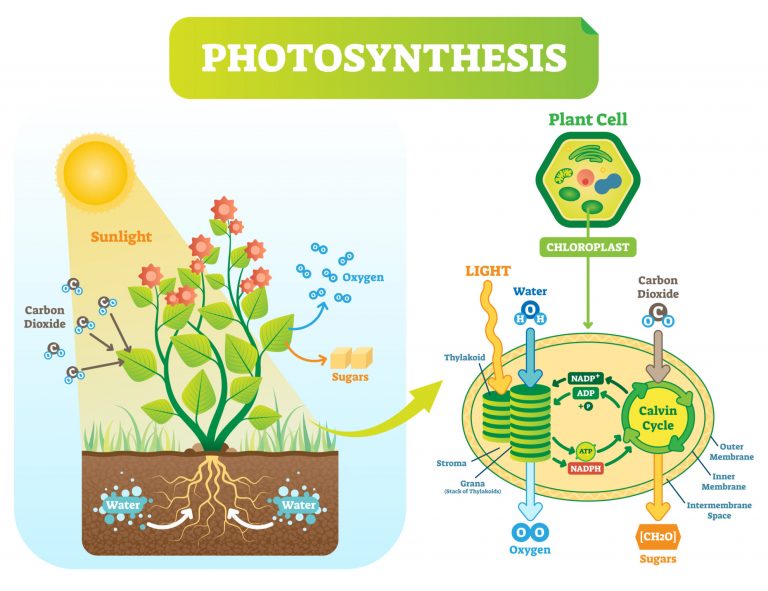
-
Saccharification
The process wherein complex carbohydrates are degraded into simpler forms, such as glucose, is called saccharification. It entails hydrolysis. In humans and other higher animals, this involves enzymatic action. In the mouth, glucose-containing complex carbohydrates are broken down into simpler forms through the action of salivary amylase. In the small intestine, the digestion of complex carbohydrates is continued. Enzymes such as maltase, lactase, and sucrase break down disaccharides into monosaccharide constituents. Glucosidases are another group of enzymes that catalyze the removal of the terminal glucose from a polysaccharide comprised chiefly of long chains of glucose.
-
Digestion (Degradation)
Glucose from the digested carbohydrates is absorbed into the epithelial cells of the small intestine (particularly in the jejunum). The cells take up glucose from the intestinal lumen through the sodium ion-glucose symport system (a form of active transport mechanism that employs sodium/glucose cotransporter 1). Glucose is released into the bloodstream by allowing it to diffuse into the capillaries through facilitated diffusion. Glucose from the bloodstream enters cells through facilitated diffusion as well. Glucose transporters (GluTs) are proteins facilitating glucose transport. When inside the cell, glucose is phosphorylated to trap it inside the cell. As an effect, glucose-6-phosphate may be used in any of the following metabolic pathways: (1) glycolysis, to synthesize chemical energy, (2) glycogenesis, where glucose is brought to the liver via the vena portae to be stored as cellular glycogen, or (3) Pentose phosphate pathway to form NADPH for lipid synthesis and pentoses for nucleic acid synthesis.
-
Glycolysis
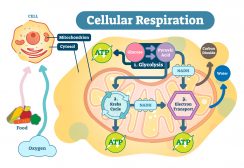
Glucose is metabolized during glycolysis. Glycolysis is a series of reactions in the cytosol that leads to the conversion of glucose into pyruvate, with the concomitant production of the relatively small amount of high-energy biomolecule, ATP. NADH, an electron-carrying molecule, is also produced. In the presence of sufficient oxygen, the pyruvate from glycolysis is metabolized further by first converting it into an organic compound inside the mitochondrion. Electron-carriers (e.g. NADH and FADH2) shuttle the electrons down the electron transport chain. A series of redox reactions occur along the chain and culminates in the final electron acceptor, i.e. the molecular oxygen. More ATP is produced via a coupling mechanism through chemiosmosis in the inner mitochondrial membrane. This three-step process is cellular respiration. From glycolysis alone, the net ATP is two (from substrate-level phosphorylation). By oxidative phosphorylation, the net ATP is about 34. Thus, the total net ATP per glucose is about 36.1
In the absence of adequate oxygen, glycolysis does not proceed to oxidative phosphorylation. Rather, anaerobic respiration ensues. Instead of shuttling the electrons in the electron transport chain, NADH transfers the electrons to pyruvate, restoring the NAD+ that sustains glycolysis.1 In anaerobic respiration, the total ATPs produced per glucose is only about two.
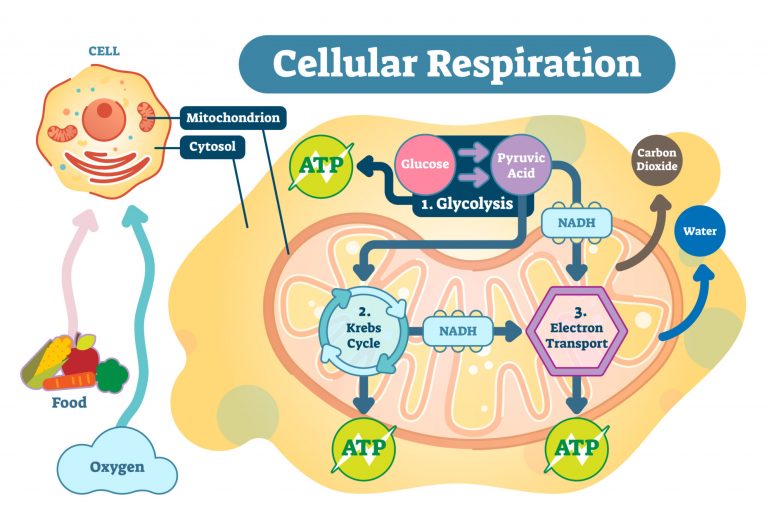
READ: Cellular respiration – Glycolysis
-
Gluconeogenesis
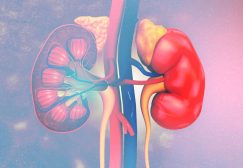
Gluconeogenesis is a metabolic process wherein glucose is generated from non-carbohydrate precursors, e.g. pyruvate, lactate, glycerol, and glucogenic amino acids. (It seems like the reverse of glycolysis in a way that glucose is converted into pyruvate whereas, in gluconeogenesis, pyruvate is converted into glucose.) The process of gluconeogenesis begins in the mitochondria and then ends in the lumen of the endoplasmic reticulum. Glucose formed from hydrolyzing glucose-6-phosphate by the enzyme glucose-6-phosphatase is shuttled from the endoplasmic reticulum into the cytoplasm. This process occurs in all organisms, including plants. In humans and many other vertebrates, it takes place mainly in the liver and the kidney and often occurs during periods of fasting, low-carbohydrate diets, or intense exercise.
-
Glycogenesis and Glycogenolysis
Glycogenesis is the metabolic process of producing glycogen from glucose for storage. It occurs chiefly in liver and muscle cells in response to the high glucose levels in the bloodstream. In the liver cell, glucose is phosphorylated by glucokinase at position 6, thus producing glucose 6-phosphate. The phosphorylation of glucose traps it inside the cell. In other cells, glucose enters passively, and then it is phosphorylated through hexokinase. This results in a compound that cannot leave the cell as well. Short polymers of glucose, especially exogenous glucose, are converted into long polymers to be stored inside the cell. The process is reversible though. When the body requires metabolic energy, glycogen is broken down into glucose subunits through the process of glycogenolysis.
In plants, starch is formed by phosphorylating glucose to produce glucose 1-phosphate, and then converting the latter into adenosine diphosphate (ADP)-glucose through the enzyme glucose-1-phosphate adenyltransferase. The ADP-glucose is added to the growing chain of glucose residues by the enzyme starch synthase. Once bound, the ADP is released and amylose is formed. In between the amylose chains, the starch branching enzyme catalyzes the formation of 1,6-alpha glycosidic bonds between the amylose chains, forming branched amylopectin.
Glycogenolysis is the process of breaking down stored glycogen in the liver so that glucose may be produced for use in energy metabolism. Thus, glycogenolysis is the opposite process of glycogenesis. Stored glycogen in the liver cells is broken down into glucose precursors. A single glucose molecule is cut off from the glycogen and is converted into glucose 1-phosphate, which in turn, is transformed into glucose 6-phosphate that can enter glycolysis. In plants, the process of breaking down stored starch is called starch degradation.
-
Pentose-Phosphate Pathway
It is a glucose metabolic pathway in which five-carbon sugars (pentoses) and NADPH are synthesized in the cytosol. It provides pentoses for use in the biosynthesis of nucleotides, such as DNA and RNA. It also synthesizes NADPH that can be used for reductive reactions during the biosynthesis of fatty acids and steroids. The pentose phosphate pathway also serves as an alternative metabolic route in the breakdown of glucose. In animals, it occurs in the liver, adrenal cortex, adipose tissues, testis, ovary, red blood cells, and mammary glands. In plants, part of the pathway functions in the formation of hexoses from carbon dioxide in photosynthesis.
-
Glycosylation
Glycosylation is the process of adding a carbohydrate component, such as glucose, to certain proteins and lipids. This step is crucial to these compounds for their proper functioning. Glycation, non-enzymatic glycosylation, is the process of covalently joining a carbohydrate constituent, such as fructose or glucose, to a protein or a lipid molecule, without the intervention of an enzyme.
Hormonal Regulation
Insulin
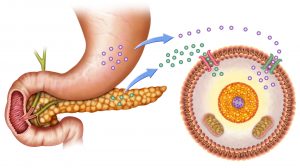
Insulin plays a role in glucose uptake in insulin-stimulated cells, such as those of adipose and muscle tissues. Insulin is released into the bloodstream when the blood glucose level is high. Insulin binds with the insulin receptor on the cell surface. The binding results in the recruitment of certain GluTs (chiefly, Glut4) located inside the cytoplasmic vesicles in the intracellular pool. The vesicles rapidly fuse into the plasma membrane where the GluTs become inserted. When the insulin stimulus is terminated, these GluTs are recycled back into the vesicles in the intracellular pool for the next insulin activation.2,3
Liver and brain cells are insulin-independent cells. These cells do not require insulin for efficient glucose uptake. Glucose is taken up by other GluTs (e.g. GluT2) that do not need a prior stimulation of insulin. This is particularly important in brain cells that always require glucose for energy metabolism. Even without insulin, glucose can essentially be taken up. Muscle cells and adipocytes, in contrast, cannot take up glucose efficiently without insulin, and therefore the lack of insulin due to low blood glucose levels will utilize alternative energy sources, such as fructose and fatty acids. Liver cells may not require insulin for glucose uptake. However, insulin still has effects on the liver. Insulin activates the enzyme hexokinase that phosphorylates glucose in order to trap it within the cell. It also activates certain enzymes involved in glycogen synthesis, e.g. phosphofructokinase and glycogen synthase. Thus, insulin tells the liver to convert glucose into glycogen by glycogenesis.3
Glucagon
Glucagon is another hormone released into the bloodstream by the pancreas when blood glucose becomes low. This hormone acts by increasing the amount of glucose in the blood. It does so by activating the enzymes involved in glycogenolysis and gluconeogenesis in the liver. It tells the hepatocytes to depolymerize glycogen to release glucose. It also stimulates the hepatocytes to use non-hexose substrates to convert them into glucose. Glucagon may also trigger lipolysis of triglycerides in the adipose tissues to provide fatty acids that can be used as an alternative energy fuel for most cells.4
Metabolic Disorders Involving Glucose
Improper metabolism of fructose results in metabolic disorders. Common metabolic disorders involving glucose may cause impaired glycolysis, glycogen synthesis, and glycogen degradation. For instance, glucose-6-phosphate isomerase deficiency affects glycolysis as the enzyme glucose-6-phosphate isomerase is used in the cytosol to convert glucose-6-phosphate to fructose-6-phosphate. The deficiency of this enzyme may be caused by a defective GPI gene on chromosome 19 in humans. Another is pyruvate kinase deficiency. Without the enzyme pyruvate kinase is deficient, glycolysis is also impaired since it is the enzyme that transfers the phosphate group from phosphoenolpyruvate to adenosine diphosphate to produce ATP and pyruvate during the last step of glycolysis. Pyruvate kinase deficiency is a heritable metabolic disorder caused by a mutation in the PKLR gene.
Other examples of inborn errors of glucose metabolism are:
- Hyperinsulinemic hypoglycemia, i.e. hyperglycemia from hyperinsulinism, due to a deficiency in glucokinase (enzyme essential in glycolysis), caused by defective GCK gene in the pancreatic beta cells
- Maturity onset diabetes of the young type II, i.e. a type of diabetes characterized by hyperglycemia from hypoinsulinemia while fasting, due to glucokinase deficiency (enzyme essential in glycolysis), caused by defective GCK gene in the pancreatic beta cells
- Glucose-6-phosphate isomerase deficiency, i.e. hemolytic anemia due to a deficiency in glucophosphate isomerase (enzyme essential in glycolysis), caused by defective GPI gene in red blood cells
- Glycogen storage disease type VII (or Tarui’s disease), i.e. a deficiency in phosphofructokinase (enzyme essential in glycolysis), caused by defective PFKL gene in the liver cell or by PFKM gene in the muscle cell
- Glycogen storage disease type XII, i.e. an aldolase A deficiency caused by a defective ALDOA gene in the muscle, liver, and red blood cells; aldolase A is an enzyme essential in glycolysis
- Triosephosphate isomerase deficiency, i.e. a deficiency in the triosephosphate isomerase (enzyme essential in glycolysis), caused by a defective TPI1 gene in the red blood cells
- GSD type X, i.e. phosphoglycerate mutase deficiency resulting in myopathy, caused by a defective PGAM2 gene in the muscle; phosphoglycerate mutase is an enzyme essential in glycolysis
- Enolase deficiency, i.e. an autoimmune disorder due to lack of sufficient enolase (enzyme essential in glycolysis), caused by a defective ENO1 gene in the red blood cells
- Pyruvate kinase deficiency, i.e. a deficiency in pyruvate kinase (enzyme essential in glycolysis), caused by a defective PKLR gene in the red blood cells and liver
- Baker-Winegrad disease, i.e. deficiency in fructose bisphosphatase (enzyme essential in gluconeogenesis), caused by a defective FBP1 gene in the liver
Biological Importance/Functions
Glucose is one of the products of photosynthesis in plants and other photosynthetic organisms. In plants, glucose molecules are stored as repeating units of sugar (e.g. starch). It is also an important component of amylopectin and cellulose. Thus, it occurs abundantly in fruits, plant juices, and many other plant organs.
Glucose also serves as an important metabolic intermediate of cellular respiration. It is a major source of energy via aerobic respiration, anaerobic respiration, or fermentation. In animals, it circulates in the blood and as such referred to as blood sugar. Glucose molecules are transported to various parts of the body. In the liver and muscle cells, excess glucose molecules are stored as glycogen. The brain takes up glucose since it prefers glucose over the others as a source of energy. Without an adequate amount of glucose, the brain cannot function properly and processes such as decision-making and self-control are impaired. Glucose is a major source of energy not just in the brain but in the other parts of the body as well. It provides about 4 calories per gram. Thus, a dietary glucose serving of, for instance, 100 grams would be about 400 calories. Too much glucose, though, is implicated in certain diseases such as diabetes mellitus and in conditions such as hyperglycemia, glucosuria, and obesity.
Glucose is also an important constituent of several other biological compounds, such as the structural carbohydrates: cellulose and chitin. It is also a vital component of important disaccharides, such as lactose (milk sugar), maltose (malt sugar), and sucrose (table sugar). Glucose can also be broken down further and then converted into lipids. It also serves as a precursor for the synthesis of ascorbic acid. It is also used via the pentose phosphate pathway to provide pentoses for use in the biosynthesis of DNA and RNA.
Try to answer the quiz below to check what you have learned so far about glucose.

Related terms
See also
- Glycolysis
- Gluconeogenesis
- Glycogenesis
- Glycogenolysis
- Pentose phosphate pathway
- Glycosylation
- Galactose
- Fructose
- Insulin
- Diabetes mellitus
Reference
- Campbell, N. A. (1996). Biology. California: The Benjamin/Cumming Publishing Company, Inc. pp. 159-ff.
- Regulation of Glucose Uptake. (2002, July 19). Retrieved from Medscape.org/viewarticle/438374 Link
- Bowen, R. (2019). Physiologic Effects of Insulin. Retrieved from Vivo.colostate.edu/hbooks/pathphys/endocrine/pancreas/insulin-phys.html Link
- Bowen, R. (n.d.). Glucagon. Retrieved from Vivo.colostate.edu/hbooks/pathphys/endocrine/pancreas/glucagon.html Link
© Biology Online. Content provided and moderated by Biology Online Editors.