
Bondok’s Classification of the Thalamic Nuclei
Anatomists usually classify the thalamic nuclei according to their relation to the internal medullary lamina i..
Mendel’s laws: definitions
Genetics – a branch of biology that deals with the study of heredity and variations Variations – d..
Gorillas in our Midst – what it means
Have you tried gorilla on a basketball court perception test? Do you know what it implicates? Find out in this..

Public Engagement: Rising Expectations in the Digital Age
Gone are the simpler days where scientists could isolate and focus solely on their work, engaging exclusively ..
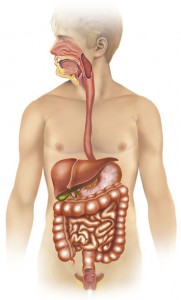
How Celiac Disease Affects The Digestive System
Celiac disease is a condition that affects the digestive systems of many individuals all over the world, and i..

CRISPR DIY – biohacking genes at home
Have you ever thought of changing yourself for the better — genetically-speaking? Lately, CRISPR company..
![Hallucinations – a brain glitch – apparently could occur in animals, too. At least, according to a recent experiment on lab mice using optogenetics technique. [Img credit: Rick Harris (Flickr), by CC BY-SA 2.0]](https://www.biologyonline.com/wp-content/uploads/2019/07/hallucination-300x168.jpg)
Hallucination – Are we the only ones “seeing” things or animals hallucinate, too?
Hallucination is defined as perceiving something that seems real but in fact, it is not. Some references take ..

A New Theory on the Origin of Animal Multicellularity
Multicellular life, purportedly, started around 600 million years ago. From single-celled, certain organisms e..
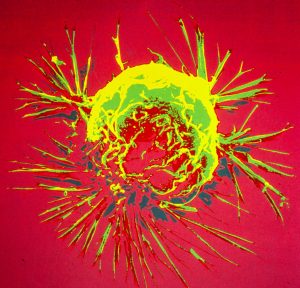
RASER proteins selectively “hack” and “shut down” cancer cells
According to World Health Organization, cancer is the second leading cause of death worldwide. The record show..
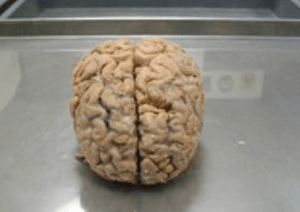
Scientists brought dead pig brain partly back to life
Death is inevitable to any entity that has life. When there is a beginning there ought to be an end. However..

RNA-DNA World Hypothesis?
How did life start as we know it? In the scientific community, the “RNA World Hypothesis“ has many..

FAAH-OUT mutation for a life of no pain – No FAAH, no pain
A Scottish woman claims that she has not experienced pain over some supposedly painful conditions, like a seve..

Regeneration in humans – Finding the gene switch
Regeneration in humans is much more limited compared in other animals. Say for instance when one lost a limb, ..

On Mate Selection Evolution: Are intelligent males more attractive?
A study published in Science on January 11 seems to be the first to lay empirical evidence that concur with Ch..

Blindness – Evolutionary regression? Maybe not!
The recent Netflix’s hit flick, Bird Box, surely startled the viewers with the thrilling scenarios revol..

Mitochondrial DNA not just from moms but also from dads?
If one wants to trace down lineage, that person could turn to the cell’s powerhouse, the mitochondrion. ..