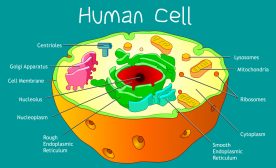
Figure
1. The form of anything; shape; outline; appearance. Flowers have all exquisite figures. (Bacon)
2. The representation of any form, as by drawing, painting, modeling, carving, embroidering, etc.; especially, a representation of the human body; as, a figure in bronze; a figure cut in marble. A coin that bears the figure of an angel. (Shak)
3. A pattern in cloth, paper, or other manufactured article; a design wrought out in a fabric; as, the muslin was of a pretty figure.
4. (Science: geometry) a diagram or drawing; made to represent a magnitude or the relation of two or more magnitudes; a surface or space inclosed on all sides; called superficial when inclosed by lines, and solid when inclosed by surface; any arrangement made up of points, lines, angles, surfaces, etc.
5. The appearance or impression made by the conduct or carrer of a person; as, a sorry figure. I made some figure there. (Dryden) Gentlemen of the best figure in the county. (Blackstone)
6. Distinguished appearance; magnificence; conspicuous representation; splendor; show. That he may live in figure and indulgence. (Law)
7. A character or symbol representing a number; a numeral; a digit; as, 1, 2,3, etc.
8. Value, as expressed in numbers; price; as, the goods are estimated or sold at a low figure. With nineteen thousand a year at the very lowest figure. (Thackeray)
9. A person, thing, or action, conceived of as analogous to another person, t
1000
hing, or action, of which it thus becomes a type or representative. Who is the figure of him that was to come. (Rom. V. 14)
10. A mode of expressing abstract or immaterial ideas by words which suggest pictures or images from the physical world; pictorial language; a trope; hence, any deviation from the plainest form of statement. To represent the imagination under the figure of a wing. (Macaulay)
11. (Science: logic) The form of a syllogism with respect to the relative position of the middle term.
12. Any one of the several regular steps or movements made by a dancer.
13. (Science: astronomy) a horoscope; the diagram of the aspects of the astrological houses.
14. Any short succession of notes, either as melody or as a group of chords, which produce a single complete and distinct impression.
a form of melody or accompaniment kept up through a strain or passage; a musical or motive; a florid embellishment.
figures are often written upon the staff in music to denote the kind of measure. They are usually in the form of a fraction, the upper figure showing how many notes of the kind indicated by the lower are contained in one measure or bar. Thus, 2/4 signifies that the measure contains two quarter notes. The following are the principal figures used for this purpose: 2/2 2/4 2/8 4/2 2/4 4/8 3/2 3/4 3/8 6/4 6/4 6/8 academy figure, Canceled figures, lay figure, etc. See academy, Cancel, lay, etc. Figure caster, or figure flinger, an astrologer. This figure caster.” figure flinging, the practice of astrology. Figure painting, a picture of the human figure, or the act or art of depicting the human figure.
(Science: chemical) figure stone, agalmatolite. Figure weaving, the art or process of weaving figured fabrics. To cut a figure, to make a display.
Origin: f, figure, L. Figura; akin to fingere to form, shape, feign. See feign.
Dictionary > Figures