Plant Metabolism

Photosynthesis and respiration as examples of plant metabolic activities
Table of Contents
Introduction
Plants are responsible for incredible feats of molecular transformation. The processes are always being studied, but there are a few basic things that are well understood at this point in history. We will be looking at photosynthesis and respiration in some detail, as always, if you have additional questions please post them on the forum.
Photosynthesis
Photosynthesis is the process by which light energy is captured, converted and stored in a simple sugar molecule. This process occurs in chloroplasts and other parts of green organisms. It is a backbone process, in the sense that all life on earth depends on it’s functioning. The following equation sums up the process:
6CO2 (carbon dioxide) + 12 H2O (water) + light energy -> C6H12O6 (glucose) + 6O2 (oxygen) +6H2O (water)
As you see from the equation, this process is vital to us as humans, because it transforms carbon dioxide into oxygen—which we enjoy with every breath!
Carbon Dioxide (CO2)
The earth’s atmosphere contains approximately 79% nitrogen, 20% oxygen, and the remaining 1% is a mixture of less common gases—including 0.039% carbon dioxide. Carbon dioxide in the atmosphere reaches plant mesophyll via the stomata. The carbon dioxide dissolves on the thin film of water that covers the outside of cells. The carbon dioxide then diffuses through the cell wall into the cytoplasm in order to reach the chloroplasts. The oceans hold a large reservoir of carbon dioxide, which keeps the atmospheric levels essentially constant. Although there are some indicators that the atmospheric levels of CO2 are rising and adding to the global warming issue. That is a whole other topic though.
Water
Water is plentiful on earth, however, it may or may not be plentiful at the location of each individual plant. Therefore, plants will close their stomata, if need be, which reduces the CO2 supply to the mesophyll. Not even 1% of the water that is absorbed by plants is used in photosynthesis, the remainder is either transpired or incorporated into protoplasm, vacuoles or other cell materials. The water utilized in photosynthesis is the source of oxygen released as a photosynthetic byproduct.
Light
Light has a dual nature, in that it exhibits properties of both waves and particles. The energy from the sun comes to earth in various wavelengths, the longest being radio waves and the shortest are gamma rays. Approximately 40% of the radiant energy the earth receives from the sun is visible light. Visible light ranges from red, 780 nanometers to violet, 390 nanometers. The violet to blue and red to orange ranges are the most often used in photosynthesis. Most light in the green range is reflected. Of the visible light that reaches a leaf, approximately 80% is absorbed. Light intensity varies widely. Time of day, temperature, season of year, altitude, latitude, and other atmospheric conditions all play roles in the intensity of the radiant energy that will reach the earth and its organisms. High-intensity light isn’t necessarily a beneficial thing for plants. In high-intensity light, photorespiration may occur, which is a type of respiration that uses oxygen and releases carbon dioxide but differs from standard aerobic respiration in the pathways that it utilizes.
Chlorophyll
A few things to know about chlorophyll before we get into the nitty-gritty of photosynthesis and respiration. There is more than one type of chlorophyll, however, they all have one atom of magnesium in the center. In some ways, the chlorophyll is quite analogous to the heme structure in hemoglobin (the iron-containing pigment that carries oxygen in the blood). Chlorophyll has a long lipid tail that anchors the molecule in the lipid layers of the thylakoid membranes—recall that thylakoids are coin-like discs in stacks within the stroma of the chloroplasts. The chloroplasts of most plants contain two types of chlorophyll embedded in the thylakoid membranes. The formula for bluish-green chlorophyll a is C55H72O5N4Mg and the formula for yellow-green chlorophyll b is C55H70O6N4Mg. In general, most chloroplast has about three times as much chlorophyll a than b. The main role of chlorophyll b is to broaden the spectrum of light available for photosynthesis: chlorophyll b absorbs light energy and transfers the energy to a chlorophyll a molecule. Other pigments are contained in chlorophyll c, d, and e and take the place for chlorophyll b in some cases. Note that all the chlorophyll molecules are related to each other and differ only slightly in molecular structure. Light-harvesting complexes contain 250 to 400 pigment molecules and are referred to as a photosynthetic unit. There are countless numbers of these units spread throughout the grana of a chloroplast. In the chloroplasts of green plants, two types of these harvesting units operate together in order to bring about the first phase of photosynthesis.
The photosynthetic process occurs in two successive processes: the light reactions and the carbon-fixing reactions.
Recommended reading: Müller‐Schüssele, S. J., et al. (2020). Chloroplasts require glutathione reductase to balance reactive oxygen species and maintain efficient photosynthesis. The Plant Journal. https://doi.org/10.1111/tpj.14791.
Read more articles like this here: The Plant Journal – provides a dynamic forum for the ever-growing international plant sciences research community and publishes in all key areas of plant biology.
The light reactions
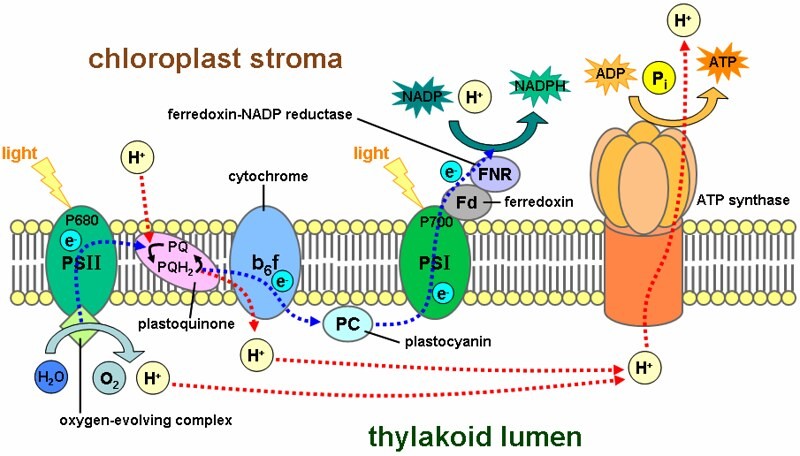
The light reactions involve light striking the chlorophyll molecules embedded in the thylakoids of chloroplasts. The subsequent reaction results in the conversion of some light energy to chemical energy. In the light reactions, water molecules are split apart into hydrogen ions and electrons and oxygen gas is released. In addition, ATP (adenosine triphosphate) molecules are created and the hydrogen ions derived from the water molecules are involved in “loading” NADP which carries the hydrogen as NADPH. NADPH is integral in providing the hydrogen ions used in the second series of major photosynthetic reactions: the carbon-fixing reactions.
The carbon-fixing reactions
The carbon-fixing reactions used to be called dark reactions because light does not play a direct role in their functioning. The reactions take place in series outside of the grana in the stroma of the chloroplast. These reactions only occur if the end products of the light reactions are available for use. Depending on the plant involved, the carbon-fixing reactions may develop or progress in different ways. The most common type of carbon-fixing reactions in plants is the process called the Calvin cycle. In the Calvin cycle, carbon dioxide from the atmosphere is combined with a 5-carbon sugar—RuBP, or ribulose bisphosphate. The combined molecules are converted via several steps into a 6-carbon sugar, such as glucose. The ATP and NADPH molecules from the light reactions provide the energy and resources for the reactions. Some of the sugars produced are further combined into polysaccharides (strings of simple sugars) or are stored as starch within the plant. There are other variations, including the 4-carbon pathway which is usually found in desert plants (C4 plants).
Before getting into respiration let’s take a closer look at what happens in both the light reactions and the carbon-fixing reactions.
Nitty-gritty of Light Reactions
Einstein called the discrete particles of light photons. Particles (photons) and waves are both currently accepted aspects of light. The quantum (energy) of photons is different depending on what kind of light they are in. Longer wavelength light has lower photon energies, while light with shorter wavelengths have higher photon energies. As mentioned earlier, every pigment color has a different distinctive pattern of light absorption—called the pigment’s absorption spectrum. The energy levels of some of the pigment’s electrons are raised when the pigment absorbs light. If energy is emitted immediately upon absorption, the effect is called fluorescence. The red part of light does this characteristically, as demonstrated when chlorophyll is placed in light it will appear red. If the absorbed energy is emitted as light after a delay, then the effect is called phosphorescence. The energy may be converted to heat or stored, as in photosynthesis within chemical bonds.
Oxidation-reduction reactions
OIL RIG, a cute little mnemonic device to remember that oxidation is loss and reduction is gain. Perhaps better put, oxidation results in the net loss of an electron or electrons, while reduction results in a net gain of an electron or electrons. The electrons come from compounds within the process or donated in from previous processes. These types of chemical reactions are found scattered throughout the processes within photosynthesis and respiration.
Photosystems
The two types of photosynthetic units in most chloroplasts are what constitute photosystem I and photosystem II.
Photosystem I contains photosynthetic units with 200 or more molecules of chlorophyll a, small amounts of chlorophyll b, protein saddled carotenoid pigment and a pair of specialized reaction-center molecules of chlorophyll called P700. All pigments in a photosystem are capable of absorbing photons, however, only the reaction-center molecules can really utilize the light energy. The other pigments aren’t worthless in the system, as they act sort of like an antenna in gathering and passing light energy along to the reaction-center. Iron-sulphur complexed proteins initially receive electrons from P700 and serve as primary electron acceptors for the unit.
Photosystem II contains chlorophyll a, protein saddled beta-carotene, a small amount of chlorophyll b and special pair of reaction-center molecules of chlorophyll a otherwise called, P680. The photosystem has a primary electron acceptor called pheophytin or Pheo.
For the record, the 680 and 700 in the names of the reaction-center molecules stand for the peaks in the absorption spectra of light waves of 680 nm and 700 nm.
Photolysis
A photon of light strikes the photosystem II reaction center, the P680 molecule to be exact near the inner surface of a thylakoid membrane. The received light energy excites an electron (boosts it to a higher energy level) which is an unstable reaction and thus most of the energy is lost to heat. Up to four photons at a time can strike the P680 molecule, however, it can only accept one electron at a time. The molecule of pheophytin picks up the excited electron, which then crosses the thylakoid membrane and is passed along to another acceptor called plastoquinone or Pq near the outside surface of the thylakoid membrane. Protein Z extracts electrons from water and replaces the ones lost by the P680 molecule. Protein Z contains manganese which is required in order to split water molecules. Simultaneously, as two water molecules are split and a molecule of oxygen and four protons are produced. This enzyme-mediated water splitting process is called photolysis.
Photophosphorylation
Pq, the acceptor molecule, releases the excited electron into the care of an electron transport system that is sort of like a downhill bucket brigade. The transport system moves electrons extracted from water temporarily to a high-energy storage molecule called nicotinamide adenine dinucleotide phosphate (NADP+). NADP+ is an electron acceptor for the photosystem. The transport chain is essentially iron-containing pigments, cytochromes, a copper-containing protein called plastocyanin and other electron transferring molecules. As electrons are passed through the chain and protons are being shuffled through a coupling factor, ATP molecules are assembled from ADP and phosphate in a process called photophosphorylation.
A similar series of events occurs in photosystem I. After a photon of light strikes a P700 molecule, the resulting excited electron is passed along to an iron-sulphur molecule Fe-S which in turn passes it to another acceptor molecule ferredoxin, (Fd). The ferredoxin molecule releases the electron to a carrier molecule called flavin adenine dinucleotide (FAD) and then eventually on to NADP+. A reduction occurs and NADP+ becomes NADPH. Electrons from photosystem II and the activities of the electron transport system replace any electrons removed from the P700 molecule. Because the electrons move in one direction, the movement of electrons from water to photosystem II to photosystem I to NADP+ are said to be part of noncyclic electron flow. Any ATP that is produced is designated noncyclic phosphorylation.
It should be noted that photosystem I can operate independently of photosystem II. When this occurs, the electrons boosted from P700 reaction-center molecules (photosystem I) are passed through an intermediary acceptor molecule called P430 and then on to the electron transport chain. This is rather than to the ferredoxin and NADP+. After being passed through the electron transport chain, the electron is dumped back into the reaction-center of photosystem I. This process demonstrates cyclic electron flow and any ATP generated by cyclic electron flow is termed cyclic phosphorylation. Note, that no water molecules are split and no NADPH or oxygen is produced.
Chemiosmosis
Earlier we mentioned in passing a coupling factor. The enzyme necessary for the mediation of the splitting of water molecules is on the inside of the thylakoid membrane. As a result of this, a proton gradient forms across the membrane and the movement of these protons is thought to be a source of energy for generating ATP. The motion is thought to be similar to molecular movement during osmosis and has hence been termed chemiosmosis. As the protons move across the membrane, they are assisted in crossing by protein channels called ATPase or coupling factor. Because of the proton movement, ADP and phosphate combine which produces ATP.
The nitty-gritty of carbon-fixing reactions
Both ATP and NADPH are important products of the light reactions and both of them play roles in the synthesis of carbohydrates from atmospheric carbon dioxide. Although the carbon-fixing reactions do not require daylight, they generally are conducted during daylight hours as there is some indication that some of the enzymes required for the processes in carbon-fixing may require some level of light. These reactions take place in the stroma of the chloroplast.
Three known mechanisms of converting carbon dioxide to sugar: (1) Calvin Cycle, (2) 4-Carbon pathway, and (3) CAM photosynthesis.
The Calvin Cycle or the 3-carbon pathway—The Calvin cycle is the most common of the three mechanisms and has four main results:
- With the assistance of the enzyme rubisco (RuBP carboxylase), six molecules of atmospheric carbon dioxide combine with six molecules of ribulose 1, 5-bisphosphate (RuBP)
- The result of the first step is six unstable 6-carbon complexes, which immediately split into two 3-carbon molecules of 3-phosphoglyceric acid or 3PGA. This is the first stable compound in photosynthesis.
- NADPH and ATP from the light-reactions, supply the energy required to convert the 3PGA to 12 molecules of glyceraldehydes 3-phosphate (GA3P), which is a 3-carbon sugar phosphate complex.
- Finally, of the 12 molecules formed; 10 are restructured into six 5-carbon molecules of RuBP—the sugar that the process started with.
The sugars produced can either add to an increase in the sugar content (carbohydrate content) of the plant or they can be used in pathways that lead to the production of lipids and amino acids.
4-Carbon Pathway—C4 plants are plants that use a 4-carbon molecule called oxaloacetic acid in place of the 3-carbon 3-phosphoglyceric acid in step two of the Calvin cycle. Oxaloacetic acid is produced from a 3-carbon compound PEP—phosphoenolpyruvate and carbon dioxide. This process is enzyme-mediated and occurs in the mesophyll cells of the leaf. Some species will convert the resulting oxaloacetic acid to aspartic, malic or other acids.
Note that the acids do not substitute for 3PGA. The 4-carbon acids migrate to the bundle sheaths surrounding the vascular bundles, where they are further converted to pyruvic acid and carbon dioxide. In returning to the mesophyll cells and interacting with ATP molecules, the pyruvic acid molecules are able to produce additional PEP. In the bundle sheath cells, the carbon dioxide formed converts into 3PGA and other molecules, by combining with RuBP. The other molecules formed are similar to the other ones formed in the Calvin cycle. The C4 cycle furnishes carbon dioxide to the Calvin cycle in a more roundabout way than the C3 pathway, but there is an advantage to this extra pathway. The extra pathway greatly reduces photorespiration in C4 plants, and this is a good thing because photorespiration is in direct competition with the Calvin cycle and even takes place in the light while the Calvin cycle is functioning. During photorespiration, RuBP reacts with oxygen to create carbon dioxide; in contrast, during photosynthesis RuBP and carbon dioxide are used to form carbohydrates. C4 plants are able to pick up carbon dioxide in very low concentrations via the mesophyll cells. The Calvin cycle occurs in the bundle sheath where carbon dioxide is readily available. Because of the location, the enzyme rubisco will be in a prime spot to catalyze the reaction between RuBP and carbon dioxide, rather than oxygen. As a result, C4 plants have photosynthetic rates that are two to three times higher than C3 plants. There are a few other characteristic features of C4 plants worth noting:
C4 plants have two types of chloroplasts and an alternate pathway for using carbon dioxide. C3 plants only have one type of chloroplast and one pathway. Chloroplasts with starch grains and are large with very little grana, and sometimes none, in the bundle sheath cells. In the mesophyll, the small, but numerous chloroplasts have no starch grains and contain highly developed grana.
PEP carboxylase is found in high concentration in the mesophyll cells which permits ready conversion of carbon dioxide to carbohydrate at lower concentrations than does rubisco (in bundle sheath cells) of the Calvin cycle.
The temperature ranges for C4 plants are much higher than C3 plants that enable C4 plants to live well in conditions that would likely kill a C3 plant.
CAM photosynthesis—Crassulacean acid metabolism is a modified photosynthetic system that is somewhat similar to C4 photosynthesis in that 4-carbon compounds are produced during the carbon-fixing reactions. CAM plants accumulate malic acid in their chlorenchyma tissues at night, which is converted back to carbon dioxide during the day. In the daytime, malic acid diffuses out of the vacuoles and is converted to carbon dioxide for use in the Calvin cycle. PEP carboxylase is responsible for converting the carbon dioxide plus PEP to malic acid at night. This modification allows for a greater amount of carbon dioxide to be converted to carbohydrate during the day than would be otherwise converted given the conditions CAM plants generally grow in. CAM plants generally close their stomata during the day in order to reduce water loss. There are more than 20 families that contain CAM plants, including cacti, stonecrops, orchids, bromeliads, and many succulents growing in regions of high light intensity. There are some succulents that do not have CAM photosynthetic capabilities, as well as non-succulents that do have the ability.
Respiration
Respiration is the group of processes that utilizes the energy that is stored through the photosynthetic processes. The steps in respiration are small enzyme-mediated steps that release tiny amounts of immediately available energy, the energy released is usually stored in ATP molecules which allow for even more efficient use of an organism’s energy. Respiration occurs in the mitochondria and cytoplasm of cells.
There are several forms of respiration: aerobic—which requires oxygen, anaerobic—which occurs in the absence of oxygen, and fermentation—which also occurs in the absence of oxygen.
Aerobic respiration is the most common form of respiration and cannot be completed without oxygen gas. The controlled release of energy is the main event in aerobic respiration.
Certain types of bacteria and other organisms without oxygen gas carry on anaerobic respiration and fermentation. Compared to aerobic respiration the amount of energy released is quite small. The main difference between aerobic respiration and fermentation is in the way hydrogen is released and combined with other substances. Two very common forms of fermentation are summed up by the following equations:
(Equation 1) C6H12O6 (glucose) -> (with enzymes)-> 2C2H5OH (ethyl alcohol)+ 2CO2 (carbon dioxide) + energy (ATP)
(Equation 2) C6H12O6 (glucose) -> (with enzymes) -> 2C3H6O3 (lactic acid) + energy (ATP)
Note the first equation is particularly valuable to the brewing industry.
Major Steps in Respiration:
Glycolysis—the first step does not require oxygen gas (O2) and takes place in the cytoplasm. The glycolytic phase is subdivided into three main steps and several smaller ones. Each step is mediated by an enzyme. A small amount of energy is released and hydrogen atoms are removed from compounds derived from glucose. The main gist of the steps are:
- The glucose molecules go through several steps and become a double phosphorylated fructose molecule.
- The 6-carbon fructose molecule is split into two 3-carbon fragments, each with a phosphate, GA3P
- Hydrogen, energy, and water are removed from the GA3P molecules leaving pyruvic acid.
Glycolysis requires two molecules of ATP to get the process started. In the processes, four ATP molecules are created, with a net gain of 2 ATP molecules at the end of glycolysis. The hydrogen ions and electrons that are released are held by an acceptor molecule called NAD—nicotinamide adenine dinucleotide. The overall end products of glycolysis is: 2-ATP molecules, 2-NADH molecules, and pyruvic acid.
The next step depends on the kind of respiration involved—aerobic, true anaerobic or fermentation. In aerobic respiration (with oxygen present), the next steps are Krebs Cycle and Electron Transport Chain.
The Krebs Cycle (or citric acid cycle)—The Krebs cycle takes place in the fluid matrix of the cristae compartments of the mitochondria. It is called the citric acid cycle because of all the intermediate acids in the cycle. The pyruvic acid product of glycolysis is restructured, some of the CO2 is lost and becomes acetyl CoA which then dumps into the Krebs cycle. During the restructuring of pyruvic acid, a molecule of NADH is produced. The Krebs cycle removes energy, CO2, and hydrogen from acetyl CoA via enzyme-mediated reactions of organic acids.
The hydrogen removed during the Krebs cycle is picked up by FAD and NAD acceptor molecules. The end result of the metabolizing of two acetyl CoA molecules in the Krebs cycle is two ATP molecules, oxaloacetic acid (to further drive the cycle), six NADH2 molecules, two FADH2 molecules, and two CO2 molecules.
The NAD and FAD molecules and the hydrogens that they carry will be dumped into the next step in respiration in order to extract the energy from the molecules.
The Electron Transport Chain—The electron transport chain (ETC) is a bit like a bucket brigade in that the chain passes the molecules along until the job is done. Energy is released as the hydrogen and electrons from the NAD+ and FAD+ carrier molecules are dumped into the system. When the electrons reach the end of the chain they pick up oxygen and water is released. ATP is produced by oxidative phosphorylation during the action of the electron transport chain. This occurs essentially like chemiosmosis.
Aerobic respiration yield
As a whole, from glycolysis to finish aerobic respiration yields the following:
Glycolysis:
- 4 molecules of ATP +2 molecules of NADH (which yields 4 ATP in the ETS) = 8 molecules of ATP net
- 8 molecules of ATP net – 2 ATP molecules to start the glycolysis process = 6 ATP molecules
Conversion of pyruvic acid to acetyl CoA:
- 2 molecules of NADH (yields 6 ATP in the ETS)
Krebs Cycle:
- 2 molecules of ATP + 2 molecules of FADH2 (which yields 4-ATP in the ETS) + 6 molecules of NADH2 (which yields 18-ATP in the ETS) = Total ATP yield: 36
The 36 resulting ATP molecules represent approximately 39% of the energy in a molecule of glucose. Compared to each other, aerobic respiration is about six times as efficient as anaerobic respiration.
Anaerobic respiration and fermentation result in a net gain of 2 ATP molecules from glycolysis. It should be noted, that the by-products of these processes, lactic acid, and alcohol, will eventually kill the organism if the products are not digested.
Factors regulating the rate of respiration
Temperature—To a point, the higher the temperature the faster respiration occurs. At some temperature, enzymes will become inactivated, although there are thermophilic (heat-loving) organisms that do quite well in high-temperature environments. Energy from sugar is released faster as the rate of respiration increases which results in a net weight loss. Plants offset the weight loss by increasing photosynthetic production of sugar. Note that during respiration, some of the energy is lost as heat, which results in an overall increase in organism temperature—not necessarily detectible by human hands.
Water—Enzymes generally operate in the presence of water, and reduced water in a plant will reduce the rate of respiration. Seeds usually have a water content of less than 10%, while mature living cells usually are in excess of 90% water. Seeds keep better if they are kept dry as the respiration rate remains quite low. However, if a seed comes into contact with water and via imbibition swells, the respiration rate will skyrocket. The temperature could increase to the point of killing the seeds. Spontaneous combustion can occur from the respiration generated heat when a fungus or bacterium is permitted to grow on wet seeds. Kind of a neat little trivia fact to tuck away.
Oxygen—Oxygen is an important regulator of respiration. If oxygen is drastically reduced, respiration may drop off to the point of retarding growth or death. Low oxygen concentrations can lead to the onset of fermentation processes.
Assimilation and Digestion
Assimilation is the conversion of the sugar produced by photosynthesis to fats, proteins, complex carbohydrates, and other substances. While digestion is the breakdown of large insoluble molecules by hydrolysis to smaller soluble forms that can be transported to various parts of the plant.
Photosynthesis vs. Respiration
Summary of key differences between photosynthesis and respiration:
Photosynthesis
- Energy stored in sugar molecules
- Carbon dioxide and water used
- Increases weight
- Requires light
- Occurs in chlorophyll
- In green organisms, produces oxygen
- With light energy, produces ATP
Respiration
- Energy released from sugar molecules
- Carbon dioxide and water released
- Decreases weight
- Can occur in light or darkness
- Occurs in all living cells
- Uses oxygen (aerobic respiration)
- With energy released from sugar, produces ATP.

You will also like...
Human Reproduction
Humans are capable of only one mode of reproduction, i.e. sexual reproduction. Haploid sex cells (gametes) are produced ..

Meiosis and Alternation of Generations
Plants are characterized by having alternation of generations in their life cycles. This tutorial is a review of plant m..
Protein Variety
The sequence of amino acids determines the type of protein. Protein is synthesized according to the sequence of nucleoti..

Seed Plants
Seed plants are vascular plants. They differ from the other vascular plants in producing seeds that germinate into a new..

Plant Metabolism
Plants are responsible for incredible feats of molecular transformation. Plant processes, such as photosynthesis, photop..

Effect of Chemicals on Growth & Development in Organisms
Plants and animals need elements, such as nitrogen, phosphorus, potassium, and magnesium for proper growth and developme..