Growth and Plant Hormones

Various growth hormones are at play for plant growth
Table of Contents
Growth
All living organisms begin in the same form: as a single cell. That cell will divide and the resulting cells will continue dividing and differentiate into cells with various roles to carry out within the organism. This is life and plants are no different. Plant growth can be determinate or indeterminate, meaning some plants will have a cycle of growth, then a cessation of growth, breakdown of tissues and then death (think of a radish plant or a tomato plant) while others (think of a giant cedar tree) will grow and remain active for hundreds of years. A tomato plant is fairly predictable and is said to have determinate growth, while the cedar tree has indeterminate growing potential. Development refers to the growth and differentiation of cells into tissues, organs, and organ systems. This again all begins with a single cell.
Plant Growth Regulators and Enzymes
Genetic information directs the synthesis and development of enzymes that are critical in all metabolic processes within the plant. Most enzymes are proteins in some form or another, produced in very minute quantities, and produced on-site—meaning they are not transported from one part of the organism to another. Genetic information also regulates the production of hormones, which will be addressed shortly. The major difference is that hormones are transported from one part of the plant to another as needed. Vitamins vital in the activation of enzymes and are produced in the cytoplasm and membranes of plant cells. Animals and humans utilize plants in order to provide some vitamin resources. In general, hormone and vitamin effects are similar and are difficult to distinguish in plants, and both are referred to in general as plant growth regulators.
Recommended Source: Plant Organ Growth – Journal of Experimetal Biology (Oxford Academic). A compilation of scientific papers on plant organ growth and development.
Plant Hormones
(Image credit: Anjanim734), CC BY-SA 4.0)
The growth and development of a plant are influenced by genetic factors, external environmental factors, and chemical hormones inside the plant. Plants respond to many environmental factors such as light, gravity, water, inorganic nutrients, and temperature.
Recommended reading: Kumar, R., Khurana, A., & Sharma, A. K. (2013). Role of plant hormones and their interplay in development and ripening of fleshy fruits. Journal of Experimental Botany, 65(16), 4561–4575. https://doi.org/10.1093/jxb/eru277.
Find more articles like this here: eXtra Botany – Journal of Experimental Botany.
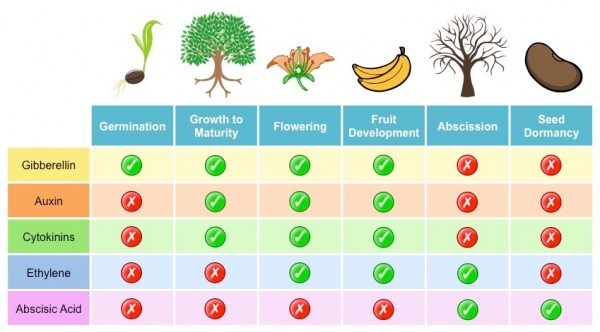
Groups of Hormones
Plant hormones are chemical messengers that affect a plant’s ability to respond to its environment. Hormones are organic compounds that are effective at very low concentrations; they are usually synthesized in one part of the plant and are transported to another location. They interact with specific target tissues to cause physiological responses, such as growth or fruit ripening. Each response is often the result of two or more hormones acting together.
Because hormones stimulate or inhibit plant growth, many botanists also refer to them as plant growth regulators. Many hormones can be synthesized in the laboratory, increasing the quantity of hormones available for commercial applications. Botanists recognize five major groups of hormones: auxins, gibberellins, ethylene, cytokinins, and abscisic acid.
Auxins
Auxins are hormones involved in plant-cell elongation, apical dominance, and rooting. A well known natural auxin is indoleacetic acid or IAA which is produced in the apical meristem of the shoot. Developing seeds produce IAA, which stimulates the development of fleshy fruit. For example, the removal of seeds from a strawberry prevents the fruit from enlarging. The application of IAA after removing the seeds causes the fruit to enlarge normally. IAA is produced in actively growing shoot tips and developing fruit, and it is involved in elongation. Before a cell can elongate, the cell wall must become less rigid so that it can expand. IAA triggers an increase in the plasticity, or stretchability, of cell walls, allowing elongation to occur.
Synthetic Auxins
Chemists have synthesized several inexpensive compounds similar in structure to IAA. Synthetic auxins, like naphthalene acetic acid, of NAA, are used extensively to promote root formation on stem and leaf cuttings. Gardeners often spray auxins on tomato plants to increase the number of fruits on each plant. When NAA is sprayed on young fruits of apple and olive trees, some of the fruits drop off so that the remaining fruits grow larger. When NAA is sprayed directly on maturing fruits, such as apples, pears, and citrus fruits, several weeks before they are ready to be picked; NAA prevents the fruits from dropping off the trees before they are mature. The fact that auxins can have opposite effects, causing fruit to drop or preventing fruit from dropping, illustrates an important point. The effects of a hormone on a plant often depend on the stage of the plant’s development.
NAA is used to prevent the undesirable sprouting of stems from the base of ornamental trees. As previously discussed, stems contain a lateral bud at the base of each leaf. In many stems, these buds fail to sprout as long as the plant’s shoot tip is still intact. The inhibition of lateral buds by the presence of the shoot tip is called apical dominance. If the shoot tip of a plant is removed, the lateral buds begin to grow. If IAA or NAA is applied to the cut tip of the stem, the lateral buds remain dormant. This adaptation is manipulated to cultivate beautiful ornamental trees. NAA is used commercially to prevent buds from sprouting on potatoes during storage.
Another important synthetic auxin is 2,4-D, which is an herbicide, or weed killer. It selectively kills dicots, such as dandelions and pigweed, without injuring monocots, such as lawn grasses and cereal crops. Given our major dependence on cereals for food; 2,4-D has been of great value to agriculture. A mixture of 2, 4-D, and another auxin, called Agent Orange, was used to destroy foliage in the jungles of Vietnam. A non-auxin contaminant in Agent Orange has caused severe health problems in many people who were exposed to it.
Gibberellins
In the 1920s, scientists in Japan discovered that a substance produced by the fungus Gibberella caused fungus-infected plants to grow abnormally tall. The substance, named gibberellin, was later found to be produced in small quantities by plants themselves. It has many effects on a plant but primarily stimulates elongation growth. Spraying a plant with gibberellins will usually cause the plant to grow to a larger than expected height, i.e. greater than normal.
Like auxins, gibberellins are a class of hormones that have important commercial applications. Almost all seedless grapes are sprayed with gibberellins to increase the size of the fruit and the distance between fruits on the stems. Beer makers use gibberellins to increase the alcohol content of beer by increasing the amount of sugar produced in the malting process. Gibberellins are also used to treat seeds of some food crops because they will break seed dormancy and promote uniform germination.
Ethylene
The hormone ethylene is responsible for the ripening of fruits. Unlike the other four classes of plant hormones, ethylene is a gas at room temperature. Ethylene gas diffuses easily through the air from one plant to another. The saying “One bad apple spoils the barrel” has its basis in the effects of ethylene gas. One rotting apple will produce ethylene gas, which stimulates nearby apples to ripen and eventually spoil because of over-ripening.
Ethylene is usually applied in a solution of ethephon, a synthetic chemical that breaks down and releases ethylene gas. It is used to ripen bananas, honeydew melons, and tomatoes. Oranges, lemons, and grapefruits often remain green when they are ripe. Although the fruit tastes good, consumers often will not buy them, because oranges are supposed to be orange, right? The application of ethylene to green citrus fruit causes the development of desirable citrus colors, such as orange and yellow. In some plant species, ethylene promotes abscission, which is the detachment of leaves, flowers, or fruits from a plant. Cherries and walnuts are harvested with mechanical tree shakers. Ethylene treatment increases the number of fruits that fall to the ground when the trees are shaken. Leaf abscission is also an adaptive advantage for the plant. Dead, damaged or infected leaves drop to the ground rather than shading healthy leaves or spreading disease. The plant can minimize water loss in the winter when the water in the plant is often frozen.
Cytokinins
Cytokinins promote cell division in plants. Produced in the developing shoots, roots, fruits, and seeds of a plant, cytokinins are very important in the culturing of plant tissues in the laboratory. A high ratio of auxins to cytokinins in a tissue-culture medium stimulates root formation. A low ratio promotes shoot formation. Cytokinins are also used to promote lateral bud growth in flowering plants.
Abscisic Acid
Abscisic acid, or ABA, generally inhibits other hormones, such as the auxin IAA. It was originally thought to promote abscission, hence its name. Botanists now know that ethylene in the main abscission hormone. ABA helps to bring about dormancy in a plant’s buds and maintains dormancy in its seeds. ABA causes the closure of a plant’s stomata in response to drought. Water stressed leaves produce large amounts of ABA, which triggers potassium ions to be transported out of the guard cells. This causes stomata to close, and water is held in the leaf. It is too costly to synthesize ABA for commercial agriculture use.
Other Growth Regulators
Many growth regulators are widely used on ornamental plants. These substances do not fit into any of the five classes of hormones. For example, utility companies all over the country often apply growth retardants, chemicals that prevent plant growth, to trees in order to prevent them from interfering with overhead utility lines. It is less expensive to apply these chemicals than to prune the trees, not to mention safer for the utility workers. Also, azalea growers sometimes apply a chemical to the terminal buds rather than hand-pruning them. Scientists are still searching for a hormone to slow the growth of lawn grass so that it doesn’t have to be mowed so often.
Plant Movements
Plants appear immobile because they are usually rooted in one place. However, time-lapse photography reveals that parts of plants frequently move. Most plants move too slowly for the passerby to notice. Plants move in response to several environmental stimuli, such as light, gravity, and mechanical disturbances. These movements fall into two groups: tropisms and nastic movements.
Tropisms
A tropism is a plant movement that is determined by the direction of an environmental stimulus. The movement toward an environmental stimulus is called a positive tropism, and movement away from a stimulus is called a negative tropism. Each kind of tropism is named for its stimulus. For example, a plant movement in response to light coming from one particular direction is called phototropism. The shoot tips of a plant that grow toward the light source are positively phototropic.
Phototropism
Phototropism, as mentioned, is illustrated by the movement of sprouts in relation to a light source direction. Light causes the hormone auxin to move tot he shaded side of the shoot. The auxin causes the cells on the shaded side to elongate more than the cells on the illuminated side. As a result, the shoot bends toward the light and exhibits positive phototropism. In some plant stems, phototropism is not caused by auxin presence or movement. In these instances, light causes the production of a growth inhibitor on the illuminated side of the shoot. Negative phototropism is sometimes seen in vines that climb on flat walls where coiling tendrils have nothing to coil around. These vines have stem tips that grow away from the light or better put, toward the wall. This brings adventitious roots or adhesive discs in contact with the wall on which they can cling and climb.
Solar tracking is the motion of leaves or flowers as the follow the suns’ movement across the sky. By continuously facing toward a light source, moving or not, the plant maximizes the light available for photosynthesis.
Thigmotropism
Thigmotropism is a plant growth response to touching a solid object. Tendrils and stems of vines, such as morning glories, coil when they touch an object. Thigmotropism allows some vines to climb other plants or objects, thus increasing its chance of intercepting light for photosynthesis. It is thought that auxin and ethylene are involved in this response.
Gravitropism
Gravitropism is a plant growth response to gravity. A root usually grows downward and a stem usually grows upward; that is, roots are positively gravitropic and stems are negatively gravitropic. Like phototropism, gravitropism appears to be regulated by auxins. One hypothesis proposes that when a seedling is placed horizontally, auxins accumulate along the lower sides of the root and the stem. This concentration of auxins stimulates cell elongation along the lower side of the stem, and the stem grows upward. A similar concentration of auxins inhibits cell elongation in the lower side of the root, and thus the root grows downward.
Chemotropism
Chemotropism is a plant growth response to a chemical. After a flower is pollinated, a pollen tube grows down through the stigma and style and enters the ovule through the micropyle. The growth of the pollen tube in response to chemicals produced by the ovule is an excellent example of chemotropism.
Nastic Movements
Plant movements that occur in response to environmental stimuli, but that are independent of the direction of the stimuli are called nastic movements. These movements are regulated by changes in water pressure in certain plant cells.
Thigmonastic movements
Thigmonastic movements are a type of nastic movements that occur in response to touching or shaking a plant. Many thigmonasties involve rapid plant movements, such as the closing of the leaf trap of a Venus flytrap plant or the folding of a plant’s leaves in response to being touched. Some leaves of sensitive plants will fold within a few seconds after being touched. This movement is caused by the rapid loss of turgor pressure (water pressure) in certain cells, a process similar to that which occurs in guard cells in order to close stomata. Physical stimulation of the plant leaf causes potassium ions to be pumped out of the cells at the base of the leaflets and petioles. Water then moves out of the cells by osmosis. As the cells shrink, the plant leaves move. It is believed that the folding of a plant’s leaves in response to touch is to discourage insect feeding.
In addition, thigmonastic movements help prevent water loss in plants. When the wind blows across a plant, the rate of transpiration is increased. If the leaves of a plant fold in response to the “touch” of the wind, water loss is reduced.
Nyctinastic movements
Nyctinastic movements are plant movements in response to the daily cycle of light and dark. Nyctinastic movements involve the same type of osmotic mechanism as thigmonastic movements, but the changes in turgor pressure are more gradual. Nyctinastic movements occur in many plants. Examples of plants that demonstrate these movements include honeylocust trees, silk trees, and bean plants. The prayer plant gets its name from the fact that its leaf blades are vertical at night, resembling praying hands. During the day, however, the leaf blades are positioned horizontally. Carolus Linnaeus planted a “flower clock” made of different species of plants with nyctinastic movements. The movements of each plant species occurred at a specific time of day when the light was right for the plant.
Seasonal Responses
In nontropical areas, plant responses are strongly influenced by seasonal changes. For example, many trees shed their leaves in the fall, and most plants flower only at certain times of the year. Plants are able to sense seasonal changes. Although temperature changes are involved in some cases and to certain degrees, plants mark the seasons primarily by sensing changes in night length.
Photoperiodism
A plant’s response to changes in the length of days and nights is called photoperiodism. Photoperiodism affects many plant processes, including the formation of storage organs and bud dormancy. However, the most studied photoperiodic process is flowering. Some plants require a particular night length to flower. In other species, a particular night length merely makes a plant flower sooner than it otherwise would.
Critical Night Length
It has been discovered that the important factor in flowering is the amount of darkness, or night length, that a plant receives. Each plant species has its own specific requirements for darkness, called the critical night length. Although it is now understood that night length, and not day length, regulates flowering, the terms short-day plant and long-day plant are still used. A short-day plant flowers when the days are short and the nights are long. Conversely, a long-day plant flowers when the days are long and the nights are short compared to the requirements of another plant.
Responding to day length and night length
Plants can be divided into three groups, depending on their response tot he photoperiod, which again acts a season indicator.
One group, called day-neutral plants (DNPs) are not affected by day length. Examples of DNPs for flowering include tomatoes, dandelions, roses, corn, cotton, and beans.
Short-day plants (SDPs) flower in the spring or fall, when the day length is short. For example ragweed flowers when the days are shorter than 14 hours and poinsettias flower when the days are shorter than 12 hours. Chrysanthemums, goldenrods, and soybeans are SDPs for flowering.
Long-day plants (LDPs) flower when the days are long, usually in summer. For example, wheat flowers only when the days are longer than 10 hours. Radishes, asters, petunias, and beets are LDPs for flowering.
Phytochrome regulation in plants
Plants monitor changes in day length with a bluish, light-sensitive protein pigment called phytochrome. Phytochrome exists in two forms, based on the wavelength of the light that it absorbs. It is generally produced in meristematic tissues in very minute amounts. The two stable forms can be converted to each other by absorbing light. Pred (Pr) that absorbs red light and Pfar-red (Pfr) that absorbs far-red light. In the daylight, more Pr is converted to Pfr (the active form) than vice versa. Pfr will convert back to Pr over several hours in the dark where it would be stable indefinitely. The conversion in light is almost instantaneous. The phytochrome mechanism is what transforms the crook in the hypocotyls of the emerging seedling into a straight stalk. Stem elongation appears to be inhibited by Pfr. However, if light levels are low, the shaded stems of a tree, for example, more far-red light will reach them and cause the conversion to Pr which lowers inhibition and allows the stems to grow longer and out from under the shade.
The interconversion abilities of phytochrome:

Vernalization
Vernalization is the low-temperature stimulation of flowering. Vernalization is important for fall-sown grain crops, such as winter wheat, barley, and rye. For example, wheat seeds are sown in the fall and survive the winter as small seedlings. Exposure to cold weather causes the plants to flower in the early spring, and an early crop is produced. If the same wheat is sown in the spring, it will take about two months longer to produce a crop. Thus, cold temperatures are not absolutely required for most crops, but they do expedite flowering. Farmers often use vernalization to grow and harvest their crops before a summer drought sets in and stunts growth.
A biennial plant is a plant that lives for two years, usually producing flowers and seeds during the second year. Biennial plants, such as carrots, beets, celery, and foxglove, survive their first winter as short plants. In the spring their flowering stem elongates rapidly, a process called bolting. Most biennials must receive cold weather to vernalize before they flower during the second year. They will then die after flowering. Treating a biennial with gibberellin is sometimes a substitute for cold temperatures in vernalization, and will stimulate the plant to grow.
Fall colors
Some tree leaves are noted for their spectacular fall color display. The changing fall colors are caused primarily by a photoperiodic response but also by a temperature response. As nights become longer in the fall, leaves stop producing chlorophyll. As the chlorophyll chemically degrades, it is not replaced. Other leaf pigments, the carotenoids, become visible and the green/orange splotches become more visible as the green chlorophyll turns orange. Carotenoids include the orange carotenes and the yellow xanthophylls. Anthocyanins produce the deep red and purplish-red colors in the fall display.

You will also like...

Physical Development in Humans
This tutorial elaborates on the physical development of humans, particularly from puberty to adulthood. Read this tutori..

Community Patterns
Learn about community patterns and the ecological factors influencing these patterns. Revisit some of the ecosystems you..

The Dinosaurs
Dinosaurs represented a major turn in the evolutionary development of organisms on Earth. The first dinosaurs were presu..

Pollution in Freshwater Ecosystems
There are many environmental factors that arise due to the usage of water in one way or another and for every action tha..

Growth Patterns
This tutorial describes the sigmoid curve, annual plant growth, tree growth, human growth, and insect growth as the grow..

Still Freshwater & Plants
Plants in lentic habitats have features not found in terrestrial plants. They acquired these features as they adapt to t..